- 移动端


艾佧科技(北京)有限公司
8 年
手机商铺
- NaN
- 0
- 1
- 0
- 3



技术资料/正文
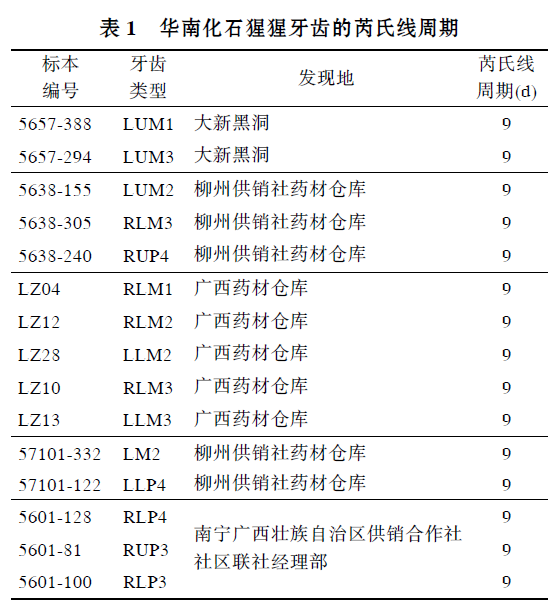
华南化石猩猩牙齿的芮氏线生长周期
864 人阅读发布时间:2022-03-08 14:43

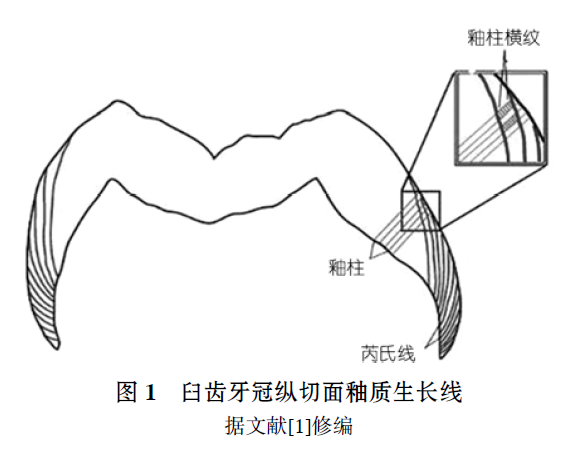
研究牙齿生长发育能为复原化石种类的个体生活历史及系统发育提供有用信息. 牙齿生长线是牙齿表面和内部的周期性发育记录, 反映了牙齿的生长发育过程. 牙组织学研究中,牙釉质生长线结构(见图1)主要包括: 釉柱横纹(Cross-striation); 芮氏线(Retzius line)及其表面特征釉面横纹(Perikymate). 釉柱横纹为短周期生长线, 生长周期为1d. 芮氏线为长周期生长线, 不同物种之间有差别, 最早由Retzius于1837年发现和描述, 是牙齿生长发育研究的重要内容[2]. 芮氏线周期是精确估计灵长类牙齿形成时间, 幼年个体年龄判定及个体发育特征的重要参数, 并具有分类学意义[3]. 最近二十多年来, 国外发表了大量的牙齿生长发育研究成果, 研究材料包括现生和化石灵长类、早期人类和现代人. 在国内, Zhao等人[4~6]曾研究了禄丰古猿和步氏巨猿的牙齿生长发育问题, 这些成果中, 确定芮氏线周期是一项重要内容, 已积累了一定量的数据结果. 但有关华南猩猩牙齿的生长发育研究仍是空白.
猩猩是亚洲地区唯一的现生大型类人猿, 生活于印尼加里曼丹和苏门答腊的热带密林, 但化石记录表明更新世时期猩猩类动物曾广泛分布于东南亚和华南地区. 最近几年华南发现更多的猩猩类化石新材料, 目前已经发现牙齿数千枚, 分布于华南30多个化石点, 时代从早更新世到晚更新世[7~9]. 这批更新世猩猩类化石涉及晚中新世以来亚洲地区大型类人猿演化、分类及早期人类起源等重要学术问题, 也是研究现生猩猩起源、演化及分类问题的重要材料. 过去, 对于华南猩猩化石的详细研究报告较少. 顾玉珉等人[9]和周国兴[7]研究了少数地点出土的猩猩化石, 并讨论了分类和演化问题. 王翠斌等人[8]通过对大量猩猩化石的测量及统计分析简单探讨了其分类问题. 赵凌霞等人[10]对广西崇左木榄山洞古人类遗址发现的猩猩化石做了较为详细的描述, 认为其有别于印尼猩猩. 不过以往所有这些研究都局限于牙齿形态学, 多将华南化石猩猩归入猩猩的一个亚种, 即猩猩魏氏亚种, 以区别于印尼猩猩. 本研究从牙齿生长发育入手, 通过制作牙组织切片, 在偏光显微镜下观察牙齿生长线结构, 分析和确定华南猩猩的牙齿芮氏线周期, 为将来进一步研究牙冠形成时间、个体发育及系统分类等内容提供依据; 同时将华南猩猩与其他古猿、早期人类、现生灵长类及现代人的芮氏线周期进行了广泛比较, 探讨不同种类之间的差异性及其系统关联; 此外, 还分析了芮氏线周期与体重的相关性, 为复原化石类人猿和人类的个体生长发育特征提供依据.
本文研究材料来自20世纪50~60年代中国科学院古脊椎动物与古人类研究所广西野外调查队在广西地区获得的的猩猩牙齿化石. 选取15枚齿冠保存完整、没有或只有轻微磨耗的猩猩牙齿化石(臼齿10 枚, 前臼齿5 枚). 其中2枚牙齿有明确出处, 来自大新黑洞地点, 地质时代为中更新世早期[11]. 5 枚来自柳州供销社药材仓库(化石主要来自来宾、柳江、柳州市及宜山、柳城等地), 3枚主要来自原南宁广西壮族自治区供销合作社区联社经理部(主要来自扶绥、崇左等县), 其他5枚也是从广西药材仓库所购, 但不能确定具体出处. 考虑到华南地区尚未发现全新世猩猩材料, 而发现的猩猩化石根据伴生动物均属更新世, 因而判断本文出处不明的猩猩牙齿化石可能也是更新世时期.
(2) 牙齿切片制作.
制作牙齿唇舌径纵向切片,并通过近中牙尖. 切片制作过程[2]: 清水超声清洁牙齿表面, 并用0.5%磷酸酸蚀牙齿表面20 s. 将颗粒状聚酯树脂铺在包埋盒底, 然后将牙齿放入盒内, 倒入液体状聚酯树脂直至浸没牙齿, 将包埋盒放入光固化仪(EXAKT 530, 德国), 蓝光及黄光照射各6 h. 将标本从包埋盒中取出, 修成合适大小. 先用微距切片机(EXAKT E300CP, 德国)沿近中唇舌径将标本剖开,然后更换为较薄的刀片, 切得厚约120~200m 牙齿薄片. 数控硬组织连续磨片机(EXAKT 400CS, 德国)磨片成80~100m, 并抛光. 最后聚酯树脂封片.
(3) 显微镜观察与芮氏线周期的确定.
在偏光显微镜(Leica DMRX, 德国)下, 首先观察确定芮氏生长线(50×). 选取釉柱横纹清晰的区域(通常在靠近釉质外表面), 在200 倍显微镜下观察计数相邻两条芮氏线之间的釉柱横纹数目, 即为芮氏线周期.
2、结果与讨论
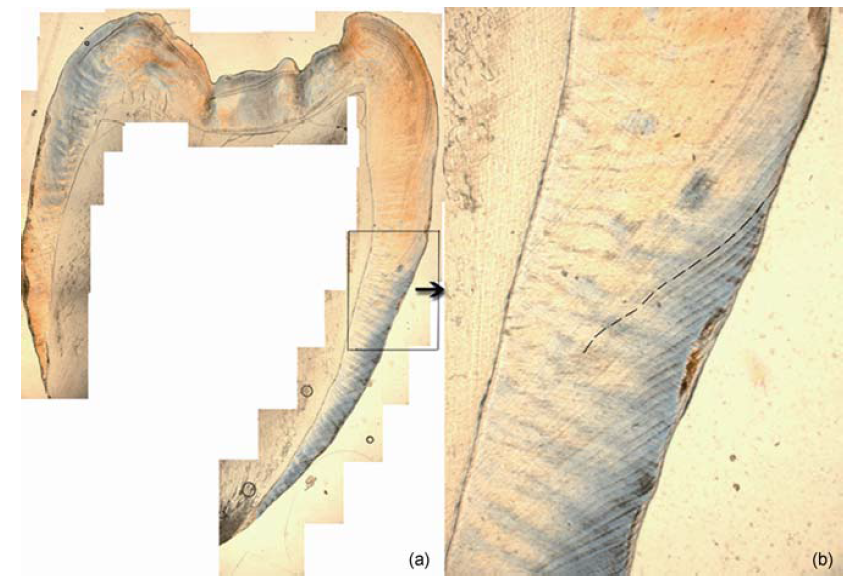
黑色虚线所示为芮氏线, 50×

图3 华南猩猩牙齿(LZ12)芮氏线及釉柱横纹
白色长线所示为芮氏线, 白色短线为釉柱横纹; 200×
2.1 与现生灵长类、现代人、古猿及早期人类的芮氏线周期比较
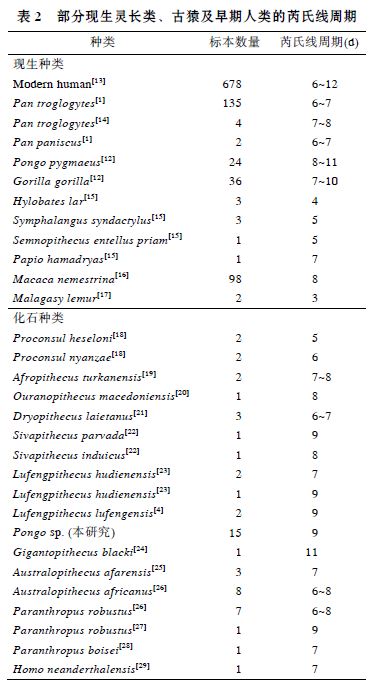
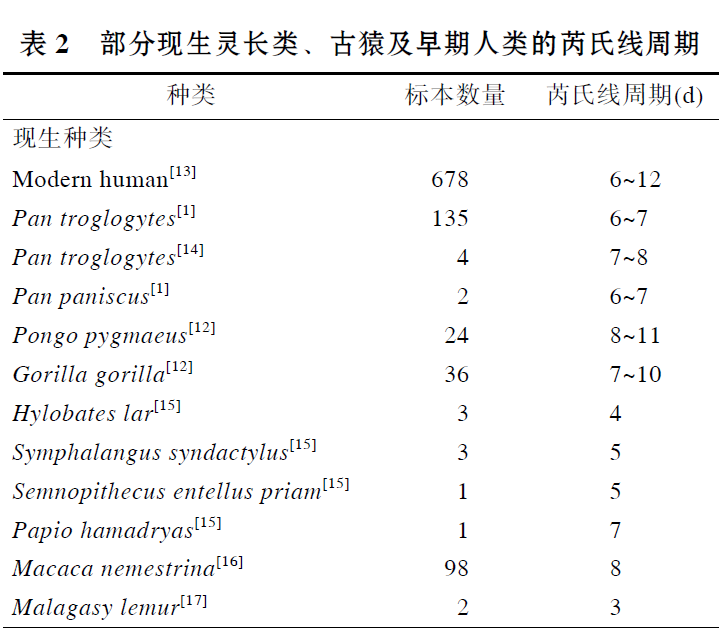
华南猩猩牙齿芮氏线生长周期为9d, 与现生灵长类比较, 在现生猩猩变异范围8~11d内[12], 也在现生大猩猩的变异范围内7~10d[12], 长于现生黑猩猩、长臂猿、合趾猿、叶猴、狒狒、猕猴以及狐猴. 华南猩猩的芮氏线周期也在现代人的变异范围 6~12d, 与现代人的平均值相[13].
与化石种类比较, 华南化石猩猩的芮氏线周期, 与禄丰古猿、西瓦古猿相当或相近, 而短于步氏巨猿; 明显长于原修康尔猿, 也长于非洲古猿、乌兰诺古猿和森林古猿. 南方古猿阿法种和非洲种、傍人粗壮种和鲍氏种等早期人类的芮氏线周期大多在 6~8d, 短于更新世华南猩猩的芮氏线周期. 整体上来看, 亚洲地区发现的大型古猿芮氏线周期似乎长于非洲和欧洲的大型古猿, 这可能与中新世以来大型猿类的辐射、扩散及地区隔离分化有关, 华南化石猩猩可能与禄丰古猿、西瓦古猿及巨猿有着更近的渊源关系.
对表2中现生灵长类的芮氏线周期比较, 发现属于原猴亚目(低等灵长类)的狐猴芮氏线周期最短, 只有3d, 显然短于猿猴亚目(高等灵长类)的三类大猿、长臂猿、合趾猿、猕猴、狒狒以及叶猴的芮氏线周期. 在猿类中, 大型猿类(猩猩、黑猩猩和大猩猩)芮氏线周期长于小型猿类(长臂猿和合趾猿); 在猴类中, 猕猴和狒狒(猕猴科)芮氏线周期长于叶猴(疣猴科). 另外, 不同地质年代的古猿芮氏线周期比较发现, 从时代最早的原修康尔猿(中新世早期)、时代较晚的几种中新世古猿到更新世的步氏巨猿, 芮氏线周期似有一个由短到长的增长趋势; 现代人的芮氏线周期变异范围较大, 其最大值显然比早期人类南方古猿的芮氏线周期要长.

2.2 芮氏线周期与体重的相关性分析
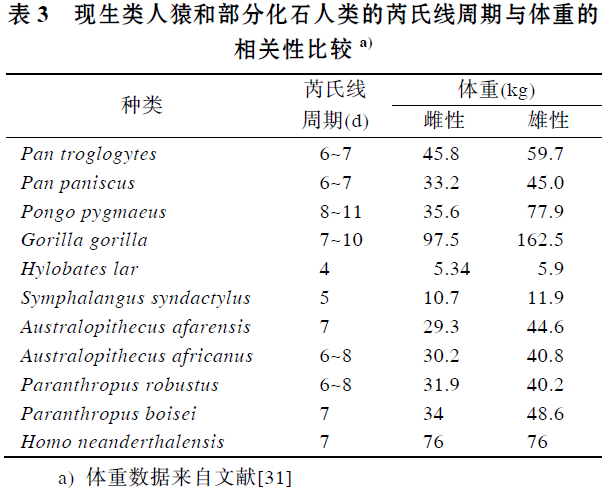
牙齿生长发育与个体生活史密切相关, 个体生活史参数包括关键的成熟事件, 例如断奶年龄、月经初潮、初次生殖的年龄、成年脑量、体重、寿命等, 而芮氏线周期是牙齿生长发育中一个非常重要的特征参数. Reid 等人[30]曾提到在类人猿中芮氏线周期体重之间存在一种正相关关系, 但没有发表详尽的研究报告. 本研究分析了6 种现生类人猿(包括普通黑猩猩、倭黑猩猩、猩猩、大猩猩、长臂猿及合趾猿)和5种化石人类(包括南方古猿非洲种、阿法种、傍人粗壮种、鲍氏种和尼安德特人)芮氏线周期和体重(见表3)之间的相关性. 在进行相关分析时, 考虑到体重的性别差异, 雌性和雄性分别比较, 体重数据均来自于文献[31], 芮氏线周期有变异的种类取其中间值. 结果发现芮氏线周期与成年体重之间呈显著正相关, 雌性相关系数为0.615(P<0.05), 雄性为0.748(P<0.01)(见图4). 推测华南化石猩猩的体重, 个体生活史特征可能在现生猩猩的变异范围内.

图4 现生类人猿及化石人类芮氏线周期与体重之间的关系
3、结论
综合上述比较讨论, 华南化石猩猩牙齿的芮氏生长周期, 与现生灵长类、化石猿类和古人类相比,属于比较长的种类; 芮氏线周期与体重之间有着显著的相关性. 相对于欧洲和非洲的中新世古猿, 亚洲地区发现的禄丰古猿、猩猩及步氏巨猿有着相对较长的芮氏线周期, 这一结果对分析不同时代和不同地区的类人猿分类、演化及生物适应性有一定的参考价值.

-
Smith T M. Incremental Development of Primate Dental Enamel. Doctor Dissertation. New York: Stony Brook University, 2004
-
Smith T M, Martin L B, Leakey M G. Enamel thickness, microstrucutre and development in Afropithecus turkanensis. J Hum Evol, 2003, 44: 286–306
-
Bromage T G, Dean M C. Re-evaluation of the age at death of immature fossil hominids. Nature, 1985, 317: 525–527
-
Zhao L X, Lu Q W, Xu Q H. Enamel microstructure of Lufengpithecus lufengensis. Acta Anthrop Sin, 2000, 19(Suppl): 77–108
-
赵凌霞, 郑良, 高峰, 等. 元谋古猿釉质微结构研究. 中国科学 D 辑: 地球科学, 2002, 32: 921–927
-
赵凌霞. 布氏巨猿牙齿的综合研究. 博士学位论文. 北京: 中国科学院研究生院, 2006
-
周国兴. 华南早期褐猿的分布与演化. 北京自然博物馆研究报告, 2002, 60: 71–87
-
王翠斌, 赵凌霞, 金昌柱, 等. 中国更新世猩猩类牙齿化石的测量研究及其分类学意义. 人类学学报, 2009, 28: 192–200
-
顾玉珉, 黄万波, 宋方义, 等. 两广部分地区猩猩化石的研究. 人类学学报, 1987, 6: 272–283
-
赵凌霞, 王翠斌, 金昌柱, 等. 广西崇左木榄山洞古人类遗址中发现的猩猩类牙齿化石及其分类演化意义. 科学通报, 2009, 54: 2920–2926
-
韩德芬. 广西大新黑洞哺乳动物化石. 古脊椎动物与古人类, 1982, 20: 59
-
Schwartz G T, Reid D J, Dean C. Developmental aspects of sexual dimorphism in hominoid canines. Int J Primatol, 2001, 22: 837–860
-
Reid D J, Dean M C. Variation in modern human enamel formation times. J Hum Evol, 2006, 50: 329–346
-
Reid D J, Schwartz G T, Dean C, et al. A histological reconstruction of dental development in the common chimpanzee, Pan troglodytes. J Hum Evol, 1998, 35: 427–448
-
Dirks W, Bowman J E. Life history theory and dental development in four species of catarrhine primates. J Hum Evol, 2007, 53: 309–320
-
Smith T M. Experimental determination of the periodicity of incremental features in enamel. J Anat, 2006, 208: 99–113
-
Schwartz G T, Godfrey L R, Mahoney P. Inferring primate growth, development and life history from dental microstructure: The case of the extinct Malagasy lemur, Megaladapis. In: Bailey S E, Hublin J-J, eds. Dental Perspectives on Human Evolution. Dordrecht: Springer, 2007. 147–162
-
Beynon A D, Dean M C, Leakey M G, et al. Comparative dental development and microstructure of Proconsul teeth from Rusinga Island, Kenya. J Hum Evol, 1998, 35: 16–209
-
Kelley J, Smith T M. Age at first molar emergence in early Miocene Afropithecus turkanensis and life-history evolution in the Homi- noidea. J Hum Evol, 2003, 44: 307–329
-
Smith T M, Martin L B, Reid D J, et al. An examination of dental development in Graecopithecus freybergi (=Ouranopithecus macedo- niensis). J Hum Evol, 2004, 46: 551–577
-
Kelley J, Dean M C, Reid D J. Molar growth in the late Miocene hominoid, Dryopithecus laietanus. In: Brook A, ed. Dental Morphology 2001: 12th International Symposium on Dental Morphology. Sheffield: Sheffield Academic Press, 2001. 123–134
-
Mahoney P, Smith T M, Schwartz G T, et al. Molar crown formation in the Late Miocene Asian hominoids, Sivapithecus parvada and Si- vapithecus indicus. J Hum Evol, 2007, 53: 61–68
-
Schwartz G T, Liu W, Zheng L. Preliminary investigation of dental microstructure in the Yuanmou hominoid (Lufengpithecus hudienen- sis), Yunnan Province, China. J Hum Evol, 2003, 44: 189–202
-
Dean M C, Schrenk F. Enamel thickness and development in a third permanent molar of Gigantopithecus blacki. J Hum Evol, 2003, 45: 381–387
-
Lacruz R S, Ramirez Rozzi F V. Molar crown development in Australopithecus afarensis. J Hum Evol, 2010, 58: 201–206
-
Lacruz R S, Ramirez Rozzi F V, Bromage T G. Variation in enamel development of South African fossil hominids. J Hum Evol, 2006, 51: 580–590
-
Dean M C. Daily rates of dentine formation in macaque tooth roots. Int J Osteoarchaeol, 1993, 3: 199–206
-
Dean M C. Growth layers and incremental markings in hard tissues: A review of the literature and some preliminary observations about enamel structure in Paranthropus boisei. J Hum Evol, 1987, 16: 157–172
-
Smith T M, Harvati K, Olejniczak A J, et al. Dental development and enamel thickness in the Lakonis Neanderthal Molar. Am J Phys Anthropol, 2009, 138: 112–118
-
Reid D J, Ferrell R, Walton P. Histological of dental derived canine crown formation times from a medieval Danish sample. Am J Phys Anthrop, 2002, 34(Suppl): 129
-
Fleagle J G. Primate Adaptation and Evolution. 2nd ed. San Diego: Academic Press, 1999
